 | Lecture 11. Competition and Cooperation |
|
F2 |
F3 - In this example, species #1 becomes extinct as a result of its competition with species #2. |
F4 - Lotka-volterra model http://home.comcast.net/~sharov/PopEcol/xls/lotkcomp.xls |
F5 |
|
F6 - This system has an unstable equilibrium which separates 2 areas of attraction: (1) where the first species ousts the second one and (2) where the second species ousts the first one -Lotka-Volterra model -http://home.comcast.net/~sharov/PopEcol/xls/compet.xls |
F7 - Cooperation. Group effect results in the increase of population growth rate with increasing population density when density is low. There is more danger of extinction in populations with group effect because there is a minimum population density below which population declines until it is gone.Cooperation between different species is relatively rare in nature and we will not study its models. Usually it has a form of symbiosis. |







Competition
Competition is among the most important factors in population dynamics of many species. It often determines the upper limit of fluctuations of population numbers. Also it determines conditions for coexistence of ecologically similar species. Intra-specific cooperation may significantly affect population dynamics at low density (or small group size).
11.1. Intra-specific competition
11.2. Competition between species
11.3. Ecological niche
11.4. Cooperation
11.1. Intra-specific competition
Intraspecific competition results in a reduction of population growth rate as population density increases. We already studied several models that consider intraspecific competition: logistic model and Ricker''s model. In these models population growth rate steadily declines with increasing population density.
However, in nature competition effect may be completely absent until population density reaches some threshold at which resources become limited F1.
Forest insect defoliators often have a "scramble" competition. If larvae can find foliage they still survive, but when all foliage is destroyed then mortality increases very rapidly. This is because these insects have seasonally-synchronized life cycles. If food is exhausted before they can pupate, then none of them pupates.
Mechanisms of competition:
1. Simultaneous resource utilization. Examples: forest defoliators, dung insects
2. Direct interaction. Examples: aggression, cannibalism, territoriality
Simultaneous resource utilization is usually associated with threshold-type relationships between resource abundance and population growth rate; in some cases "scramble" competition happens.
Direct interaction among organisms makes competition balanced and usually results in a gradual decline of population growth rate with the decrease in the amount of resources
11.2. Competition between Species
Competition among ecologically similar species is the major factor that determines the structure of animal and plant communities. The main question is, can competing species coexist or not, and what are the major factors that affect coexistence. This topic is a bridge between population ecology and community ecology.
Major problems:
1. In conservation ecology: to prevent extinction of particular species; predict potential losses in species composition after introduction of competitors; to reduce competition effects.
2. In biocontrol: to find an exotic natural enemy which will successfully fit into the community of existent natural enemies; to find exotic non-pest competitors that may oust the pest species.
In the logistic model, population density converges to the carrying capacity K, as it is shown F2
Now, we will introduce the second (competing) species. As a result, the figure becomes two-dimensional F3
Competitive exclusion principle was first formulated by Grinnell (1904) who wrote:
"Two species of approximately the same food habits are not likely to remain long evenly balanced in numbers in the same region. One will crowd out the other; the one longest exposed to local conditions, and hence best fitted, though ever so slightly, will survive, to the exclusion of any less favored would-be invader."
If competing species are ecologically identical (use the same resource), then inter-specific competition is equivalent to intra-specific competition. Each organism competes with all organisms of both populations. As a result, population growth rate of each population is determined by the sum of numbers of both populations F4
In this case, both isoclines are parallel and have a slope of 45� (see figures above). The species that have a higher carrying capacity (K) always wins. Higher carrying capacity means that the species can endure more crowding than the other species (e.g., due to more effective search for resources). Competitive exclusion is called K-selection because it always go in the direction of increasing K.
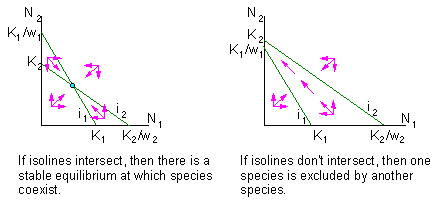
If competing species are sufficiently different then intra-specific competition is stronger than inter-specific competition. Organisms of another species are not considered as "full" competitors. As a result, the numbers of inter-specific competitors is multiplied by a weight wi<1 F5
Theoretically it is possible that weights wi>1. This means that organisms of another species are stronger competitors than organisms in the same population. I don''t know any example of this sort. But this situation is always discussed in ecological textbooks. If wi>1 and isoclines intersect, then one species will oust the second one, but what species will be excluded depends on initial conditions (initial numbers of both populations) F6
Thus, species coexistence is possible if intraspecific competition is stronger than interspecific competition. This occurs if competing species have different preferences in resource usage.
When the principle of competitive exclusion became widely known among ecologists, it seemed to contradict with some well known facts and this contradiction was formulated as "paradoxes". For example, "plankton paradox" focused on the variability of plankton organisms which all seemed to use the same resources. All plankton algae use solar energy and minerals dissolved in the water. There are not so many mineral components as compared to a large variability in plankton algae species.
There is no final solution for this paradox. However, it became clear that coexistence of species that use the same resource is a common phenomenon. Mathematical models described above are correct, but they are oversimplified; thus it is difficult to apply them to real species. More complicated and more realistic models indicate that species coexistence is possible. For example, plankton algae have distinct seasonality in their abundance which is ignored in the simple Lotka-Volterra model. Cyclical dynamic regime allows species to coexist even if they cannot coexist in stable systems. Another important factor is spatial heterogeneity which effect is substantial even in such homogeneous systems as the ocean.
11.3. Ecological Niche
The ecological niche is not a notion of quantitative population ecology despite of several attempts to define it quantitatively. There were numerous definitions of ecological niche. Grinnell (1917) defined it as all the sites where organisms of a species can live (where conditions are suitable for life). Elton (1927) described the niche as the function performed by the species in the community of which it is a member. The first definition emphasized the "address" of the species and the second one emphasized its "profession" (Miller 1967).
Hutchinson (1957) defined a niche as a region (n-dimensional hypervolume) in a multi-dimensional space of environmental factors that affect the welfare of a species. This definition is more close to Grinnell''s definition. It became popular because the range of tolerance to ecological factors can be easily measured, whereas species "profession" is hardly measurable. It is believed that the intensity of competition is proportional to the degree of niche overlapping. However, this kind of statements should be accepted with caution because: (1) measurement of niche volume is a subjective procedure, (2) some important dimensions of the niche may be not known, (3) niches change in the life-cycle, (4) niches change from one geographical region to another.
More information about niches can be found in Pielou, chapter 13
11.4. Cooperation
Intra-specific cooperation results in increased reproduction and/or survival of organisms in groups as compared to isolated organisms. This is called group effect which was first analyzed Allee (1931) and is often called "Allee effect" F7
Next - spatial dynamic
Main Text
EXELInter-Specific Competition (Lotka=Volterra Model)
Lotka-Volterra model for F6 case